

紅藻Cyanidium caldariumのPSI-LHCI超複合体の立体構造とLHCの分子進化の解明
PR TIMES / 2024年3月8日 12時45分
【研究のポイント】
・クライオ電子顕微鏡(注1)を用いた単粒子構造解析(注2)により、酸素発生型光合成(注3)を行う紅藻(注4)Cyanidium caldarium(以下、C. caldarium)の光化学系I(PSI)(注5)と集光性色素タンパク質(LHC)(注6)から構成されるPSI-LHCI超複合体の立体構造を解明しました。
・C. caldarium PSI-LHCIはPSI単量体に5個のLHCIサブユニットが結合した分子構造であることを明らかにしました。
・PSI-LHCIの構造解析に基づいたLHCIタンパク質の分子系統解析により、紅藻を起点とする紅色進化系統(注7)におけるLHCIの進化モデルを提案しました。
[画像1: https://prtimes.jp/i/96787/27/resize/d96787-27-56fec63f6c5a944e8fea-2.jpg ]
【研究概要】
静岡大学の長尾遼准教授、岡山大学の加藤公児特任准教授、沈建仁教授、理化学研究所・東北大学の米倉功治グループディレクター・教授、濱口祐准教授、京都大学の熊沢穣博士課程生、伊福健太郎教授の研究グループは、理化学研究所の堂前直ユニットリーダー、豊橋技術科学大学の広瀬侑准教授らと共に、クライオ電子顕微鏡(日本電子社 CRYO ARM 300)を用いた単粒子構造解析により、紅藻C. caldarium由来のPSI-LHCIの立体構造を1.92 Aの分解能(注8)で解明しました。得られたPSI-LHCI構造には、PSI単量体に5個のLHCIが結合していました。また、C. caldariumを含むPSI-LHCIの立体構造が明らかとなっている紅色進化系統のLHCIに焦点を絞り、分子系統解析に基づくLHCIの進化モデルを提案しました。
LHCの進化に関する研究は分子系統解析が主流ですが、この方法だけではどのLHCがPSIに結合するのかを特定することができません。本研究では、PSIに結合するLHCIを立体構造解析によって特定し、その特定されたLHCIのみに焦点を当てた分子系統解析を行いました。このアプローチにより、紅色進化系統のLHCIの分子進化について、これまでにない角度からの理解が可能になりました。このように、立体構造解析と分子系統解析を組み合わせることにより、LHCIの進化に関する研究に新たな潮流が生まれることが期待されます。
本研究成果は、2024年3月5日に、Proceedings of the National Academy of Sciences of the United States of Americaに掲載されました。
【研究者コメント】
静岡大学農学部 准教授・長尾遼(ながおりょう)
本研究では、それぞれのグループの専門的な知識や技術を融合することにより、LHCの進化に迫る新たな研究の方向性を示すことができました。今後も本研究基盤を利用することにより、LHCの進化を探求していきたいです。
【研究背景】
酸素発生型光合成は、太陽光を利用して水と二酸化炭素から有機物を合成し、副産物として酸素が発生します。シアノバクテリア、藻類、陸上植物が酸素発生型光合成を行うことにより、酸素呼吸する生物は地球上で生活できています。酸素発生型光合成を行う上で光捕集アンテナシステムは欠かせない要素です。LHCは藻類や陸上植物が持つ光捕集アンテナタンパク質で、その役割は光を捕集し、獲得したエネルギーをPSIに渡すことです。生体中でLHCはPSIに結合し、超複合体(PSI-LHCI)を形成します。しかし、LHCIのPSIへの結合箇所や発現する遺伝子が、生物種に応じて異なることが知られています。
水域に生息する藻類は、見た目の色が多様です。なかでも、紅藻を起点とする紅色進化系統の藻類は、赤、茶、黄色といった特殊な色を呈します。これは、紅色進化系統のLHCIに結合する色素分子が、陸上植物と大きく異なるためです。これまでに、紅色進化系統の4種類の藻類において、PSI-LHCIの立体構造が解明されています。しかし、紅藻C. caldariumのPSI-LHCIの立体構造は解明されていませんでした。
【研究の成果】
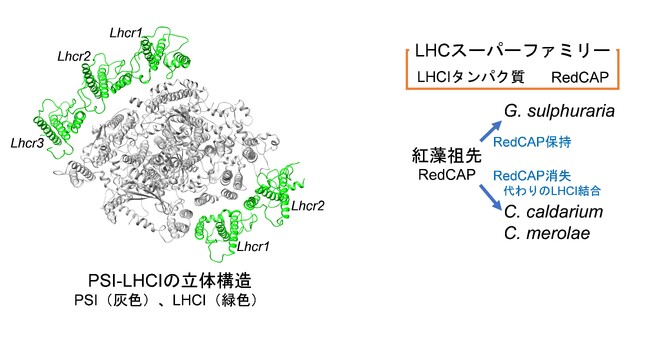
静岡大学の長尾准教授、岡山大学の加藤特任准教授、沈教授、理化学研究所・東北大学の米倉グループディレクター・教授、濱口准教授、京都大学の熊沢博士課程生、伊福教授の研究グループは、理化学研究所の堂前ユニットリーダー、豊橋技術科学大学の広瀬准教授らと共に、クライオ電子顕微鏡を用いた単粒子構造解析により、紅藻C. caldariumのPSI-LHCIの立体構造を1.92 Aの分解能で解明しました。得られたPSI-LHCI構造にはPSI単量体に3種5個のLHCIが結合しており、各LHCIをコードする遺伝子がLhcr1、Lhcr2、Lhcr3であることが明らかになりました(図1)。次に、LHCIの結合箇所と発現する遺伝子の相関を分子系統解析により調べた結果、C. caldariumのLHCIは紅藻Cyanidioschyzon merolae(以下、C. merolae)のLHCIと同じ構成であることが明らかになりました。しかし、それらの構成は、紅藻Porphyridium purpureum(以下、P. purpureum)、珪藻Chaetoceros gracilis(以下、C. gracilis)、クリプト藻Chroomonas placoidea(以下、C. placoidea)のLHCIとは大きく異なりました。
[画像2: https://prtimes.jp/i/96787/27/resize/d96787-27-2877b7e5d80db87a08c0-0.jpg ]
興味深い点として、RedCAPが挙げられます。RedCAPはLHCスーパーファミリーの中に含まれていますが、LHCIタンパク質とは進化的にかなり離れています(図2)。RedCAPは、P. purpureum、C. gracilis、C. placoideaのそれぞれのPSIには結合していますが、C. caldariumとC. merolaeには結合していませんでした。また、紅藻Galdieria sulphuraria(以下、G. sulphuraria)はゲノム中にRedCAPが存在します。C. caldarium、C. merolae、G. sulphurariaの3種類の紅藻は、原始紅藻と呼ばれており、紅藻の中でも始原的な藻類として位置づけられています。以上のことから、紅色進化系統の祖先はRedCAPを保有しているが、そこからC. caldariumとC. merolaeのRedCAPを消失させた系統とG. sulphurariaのRedCAPを保持する系統に分岐することが示唆されました。特に、RedCAPを消失したC. caldariumとC. merolaeには、RedCAPの位置に、代わりに既存のLHCIタンパク質が重複してPSIに結合するという新たな進化モデルを提案しました(図2)。
[画像3: https://prtimes.jp/i/96787/27/resize/d96787-27-71e4439c6c76d27d6ae3-1.jpg ]
【今後の展望と波及効果】
本研究によるLHCの分子系統解析は、PSI-LHCIの立体構造が解明されている5種類の紅色進化系統藻類のみで行われました。これは従来のLHCIの分子系統解析に用いられる光合成生物の数と比べると圧倒的に少ないです。LHCの遺伝子の数は生物種によって異なり、多い種だと40個以上見つかっています。しかし、すべてのLHCがPSIに結合するわけではないので、分子系統解析だけではどのLHCがPSIに結合するのかを特定することができません。本研究では、PSIに結合するLHCIを立体構造解析によって特定し、その特定されたLHCIのみに焦点を当てた分子系統解析を行いました。今後は、一つでも多くの藻類からPSI-LHCIの立体構造を明らかにし、LHCIの分子系統解析に取り組むことにより、本研究で提案した進化モデルの妥当性を検証する必要があります。このように、立体構造解析と分子系統解析を組み合わせることによりLHCの進化に迫る新たな研究手法を示すことができたため、今後のLHC研究の発展が期待されます。
【論文情報】
掲載誌名: Proceedings of the National Academy of Sciences of the United States of America
論文タイトル: The structure of PSI-LHCI from Cyanidium caldarium provides evolutionary insights into conservation and diversity of red-lineage LHCs
著者: Koji Kato, Tasuku Hamaguchi, Minoru Kumazawa, Yoshiki Nakajima, Kentaro Ifuku, Shunsuke Hirooka, Yuu Hirose, Shin-ya Miyagishima, Takehiro Suzuki, Keisuke Kawakami, Naoshi Dohmae, Koji Yonekura, Jian-Ren Shen, Ryo Nagao
DOI: https://www.pnas.org/doi/10.1073/pnas.2319658121
【用語説明】
注1:クライオ電子顕微鏡
タンパク質などの生体分子を水溶液中の生理的な環境に近い状態で、電子顕微鏡で観察するために開発された手法です。まず、試料を含む溶液を液体エタン(約-180℃)に落下させて急速凍結し、アモルファス(非晶質、ガラス状)な薄い氷に包埋します。これを液体窒素(-196℃)条件下で、透過型電子顕微鏡で観察します。電子顕微鏡内の真空中では試料は凍結状態を保持でき、また、冷却することにより電子線の照射による損傷を減らすことができます。
注2:単粒子構造解析
電子顕微鏡で撮影した多数の生体分子の像から、その立体構造を決定する構造解析手法のことをいいます。2017年のノーベル化学賞の受賞者の一人、Joachim Frankらにより単粒子解析法の基礎がつくられました。
注3:酸素発生型光合成
光合成には酸素発生型光合成と酸素非発生型光合成があります。酸素発生型光合成は、光化学系I、シトクロムb6f、光化学系II、ATP合成酵素と呼ばれるそれぞれの膜タンパク質複合体によって駆動され、光エネルギーを利用して水と二酸化炭素から炭水化物と酸素を合成します。酸素非発生型光合成生物が進化して酸素発生型光合成生物になったと考えられています。
注4:紅藻
紅藻(こうそう)とは、世界に約4,000種類いるといわれる真核藻類群です。紅藻の多くは「赤っぽい色」を呈しますが、本研究で使用したC. caldariumは緑色です。紅藻の種類は、微生物から大きな葉状構造を持つ種まで幅広く存在します。
注5:光化学系I(PSI)
光エネルギーを化学エネルギーへ変換する膜タンパク質複合体です。PSIは10種類以上のサブユニットから構成され、補欠因子として、金属錯体、色素分子(クロロフィルやカロテノイド)がタンパク質に結合しています。クロロフィルとカロテノイドはそれぞれ特有の光エネルギー吸収帯を持ち、光捕集に重要な役割を担います。
注6:集光性色素タンパク質(LHC)
太陽光エネルギーを捕集し、光化学系Iや光化学系IIに伝達するための色素タンパク質です。パラボラアンテナのように光エネルギーを集めるため、アンテナタンパク質とも言われます。集光性色素タンパク質は光合成生物種間で多様であるため、結果として、光合成生物に見た目の色の違いを生じさせています。
注7:紅色進化系統
紅藻から進化した藻類群のことで、紅藻、珪藻、褐藻、ハプト藻、渦鞭毛藻などを含みます。
注8:分解能
どのくらい細かくものを「見る」ことができるかの指標です。数値が小さい程、分解能が高いと言え、物質をより精細に観測できます。原子と原子の距離は、1.2-1.5オングストローム(A、1 Aは100億分の1メートル)程度であるので、個々の原子を区別するには1.5 A程度の空間分解能が必要となります。
企業プレスリリース詳細へ
PR TIMESトップへ

この記事に関連するニュース
-
【岡山大学】高等先鋭研究院 異分野基礎科学研究所の沈建仁所長・教授が「東レ科学技術賞」を受賞 ~光合成研究で世界を先導し、より良い社会変革を拓く~
PR TIMES / 2024年4月21日 12時15分
-
海産微細藻類における窒素固定型シアノバクテリアのオルガネラ化(細胞内小器官化)の進行を明らかに
Digital PR Platform / 2024年4月15日 14時5分
-
明大が世界に誇る研究を動画で発信!PR動画シリーズ "Incredible Senseis at Meiji University"に新しいコンテンツを公開しました ~脱炭素の新時代を作る鍵は「藻」~
Digital PR Platform / 2024年4月10日 14時5分
-
生きた細胞内で小胞の動きを可視化し、2種類の小胞融合機構を発見
Digital PR Platform / 2024年4月5日 14時0分
-
物質最薄の一次元分子ポリマーランダムネットワークの創出!~新たな触媒・分子マシン・人工知能・量子材料開発へ~
PR TIMES / 2024年4月5日 12時40分
ランキング
-
1スバル、山崎製パン、キリン……相次ぐ“事故” 問題の根っこに何がある?
ITmedia ビジネスオンライン / 2024年4月24日 8時35分
-
2蹴飛ばされて全治1ヵ月の“腰椎圧迫骨折”。貯金4,000円・借金800万円の61歳DV夫の死で、妻が決断…じつは日本で増えている「死後離婚」の壮絶な実態【行政書士が解説】
THE GOLD ONLINE(ゴールドオンライン) / 2024年4月27日 11時30分
-
3「大したことない話なので割愛します」は正しいか…75%が「割愛」を本来と違う意味で使っていることが判明
プレジデントオンライン / 2024年4月28日 8時15分
-
4明治「ザバス」が絶好調!プロテイン飲料最前線 まだ伸びるタンパク質市場へ「オイコス」も参戦
東洋経済オンライン / 2024年4月28日 7時20分
-
55月以降の日経平均上昇を裏付ける「3つの追い風」 今後もドル高円安の大幅修正は見込みづらい
東洋経済オンライン / 2024年4月28日 9時30分
記事ミッション中・・・
記事にリアクションする
![]()
記事ミッション中・・・
記事にリアクションする

エラーが発生しました
ページを再読み込みして
ください





